Bees are dying. This biological fact has become a rallying cry for some policy advocates. Different groups have raised concern over the implications of honey bee mortality—and recent increases in mortality rates—for the food supply, for the livelihoods of commercial beekeepers, for the role of pesticides in agriculture, and for the conservation of semi-wild lands and innumerable species of invertebrate pollinators. These separate issues are united by the insect most congenial to humans, Apis mellifera, the European honey bee.
The honey bee is not native to North America but was brought by European colonists in the 1600s. Managed bees moved west with American settlers, finally arriving in California in the 1850s. Bees established feral colonies early on, and managed and wild colonies of honey bees are now found across the continent.
Until the twentieth century, the primary reason for humans to keep bees was honey. As the Apis–Homo partnership evolved, the value of bees as pollinators came to be understood. Bees were valued for their contributions to (primarily) fruit production and, eventually, that value was recognized and enhanced by pollination service contracting as early as 1910. Today, U.S. markets for bees’ pollination services are routine, connecting migratory beekeepers who move their bees by truck from crop to crop—almonds, cherries, apples, pears, cranberries, blueberries, and many other fruits and vegetables. While pollinating, and after the blooming season, bees still produce honey harvested by beekeepers.
Since the beginning, American beekeepers have dealt with illnesses and parasites that afflict bees, starting with the widespread scourge of American foulbrood in the American colonies and extending through today. Arguably the most damaging threat, the invasive Varroa mite, arrived in North America from Asia in the early 1980s. But broad public awareness of honey bees and beekeeping (broad, but mostly superficial, we would argue) began in late 2006 with Colony Collapse Disorder (CCD).
In October 2006, David Hackenberg, a Pennsylvania beekeeper, took almost 3,000 honey bee colonies to Florida for the winter. In mid-November, Hackenberg discovered that two-thirds of his hives were practically empty—no adult worker bees in the hives and no dead bees nearby. Other beekeepers across the country reported similar experiences of high colony mortality and the same unusual symptoms. The phenomenon was dubbed Colony Collapse Disorder. Colonies with CCD contained brood (developing young), food stores (honey and bee pollen), and the queen—but virtually no worker bees, alive or dead.
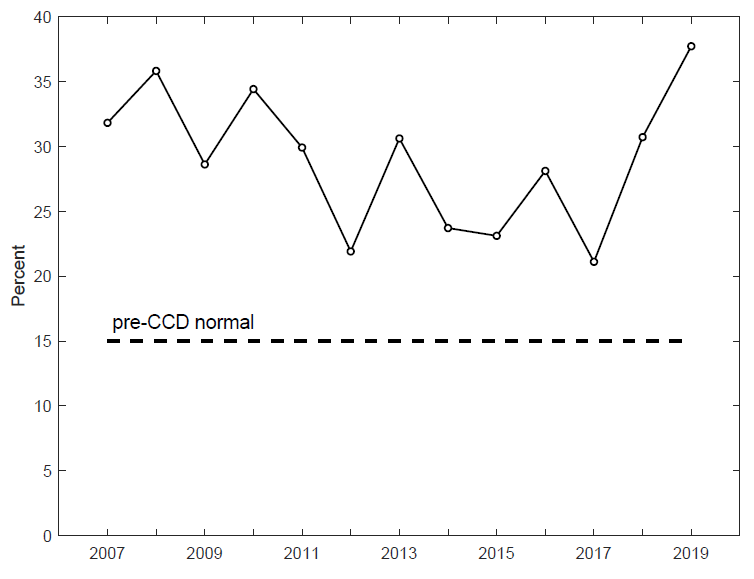
The summary indicator of CCD was an increase in overwinter mortality from a pre-CCD expectation of 15% (15 out of 100 colonies failing to survive their winter period of semi-dormancy) to rates that, since 2006 and through 2019, have averaged around 30% (see Figure 1).
Immediately following the discovery of CCD, alarm bells were rung and countless articles and reports appeared in the media. Secretary of Agriculture Mike Johanns in 2007 laid out his view of the economic threat, warning that “[i]f left unchecked, CCD has the potential to cause a $15 billion direct loss of crop production and $75 billion in indirect losses.”
Noted food writer Michael Pollan, in a 2007 New York Times article, better captured the CCD zeitgeist:
[T]he lifestyle of the modern honeybee leaves the insects so stressed out and their immune systems so compromised that, much like livestock on factory farms, they’ve become vulnerable to whatever new infectious agent happens to come along.
Press accounts of dwindling pollinators have grown steadily since that time. In 2014, President Obama, inaugurating a multiagency Pollinator Health Task Force, announced:
The continued loss of commercial honey bee colonies poses a threat to the economic stability of commercial beekeeping and pollination operations in the United States, which could have profound implications for agriculture and food (White House Office of the Press Secretary, 2014, p. 1).
In 2019, Environment America, an advocate for green issues, sent interns door to door to promote their “No Bees, No Food” campaign: “We’re here to save the bees.”
Given the saturation of public conversation with concerns about honey bee collapse, it is reasonable to ask: what is fact and what is fiction? And to ask what are the implications of bee health for the food supply?
Systematic data on beekeeper’s loss rates were not compiled prior to 2007. However, Burgett, Rucker, and Thurman (2009), Pernal (2008), and vanEngelsdorp et al. (2007) all report pre-CCD or “normal” mortality rates of about 15%, though this number is approximate and should not be taken to deny the substantial variability across beekeepers and over time. In 2007, the Apiary Inspectors of American began a telephone survey—now conducted online by the Bee Informed Partnership—compiling answers from hobbyists and commercial beekeepers. Among the questions asked of beekeepers are what percentage of their colonies failed to survive the winter. Figure 1 displays the annual averages of the response to this question along with the pre-2007, pre-CCD approximate normal level of 15% overwinter loss.
Figure 1 shows mean loss rates since 2007 to be about 30%, with considerable variability around the mean. The most recent of these values, reflecting loss rates over the winter of 2018/2019, is 38%. This is the highest loss rate reported since 2007. Whether it signals a shift in the distribution or a realization of loss rates from essentially the same distribution remains to be seen.

Figure 2 displays a different reflection of the bee economy: counts of bee colonies conducted annually by the U.S. Department of Agriculture. Data are plotted since 2000; the most recent available observation is for 2018. Given the high colony loss rates since the advent of CCD, it is perhaps surprising that the trend in colony numbers in recent years is upward (albeit moderate). In fact, the 2018 U.S. population of bees—2.8 million colonies—is higher than any observation in the past 25 years.
Figures 1 and 2 seem contradictory on their face: Colony numbers rise during the same period that overwinter mortality rates have reached historic highs. Recent work by Rucker, Thurman, and Burgett (2019) analyzes other data from the beekeeping industry (honey production, pollination fees, and queen and package bee prices) and finds similar results: little or no detectable deleterious effect after the 2007 increase in loss rates. The one possible exception to this conclusion is the fact that fees for pollinating almonds rose substantially in 2005 and 2006 (prior to CCD proper) and have plateaued at approximately $170 per hive.
The explanation for the apparent disconnect between high recent colony loss rates and the (moderate) upward trend in colony numbers is that commercial beekeepers are able to replace collapsed hives quickly and relatively inexpensively. We discuss the details of this process below.
The important point apparent in the data is that changes in colony mortality do not translate directly into changes in colony numbers. Observed colony numbers are an economic outcome, not simply a biological or environmental condition. Beekeepers and input suppliers respond to market signals in determining the number of colonies maintained (see Cheung, 1973; Rucker, Thurman, and Burgett, 2012) and, as Champetier, Sumner, and Wilen (2015) point out, colony loss and replacement result, in part, from beekeepers’ management strategies.
Honey bees have long suffered from diseases and parasites. Underwood and vanEngelsdorp (2007) document 23 episodes of major colony loss between 1868 and 2003. The most recent major predecessors to CCD are two species of mites (Varroa destructor and Acarapis woodi—tracheal mites), which first appeared in North America in the mid- to late 1980s. Varroa mites are ectoparasites that attach themselves to bees and feed on their blood (see Nordhaus, 2011, chapter 3). Tracheal mites are endoparasites that attack bees’ breathing tubes. Diseases that currently affect honey bees include American foulbrood, a bacterial infection that attacks bee larvae and pupae; Nosema, a fungus that invades the intestinal tracts of adult bees; and chalkbrood, a fungus that infests the guts of honey bee larvae. Over time, commercial beekeepers have developed methods to combat each of these parasites and diseases. That said, such methods are costly, and bee diseases and parasites have periodically devastated nonmanaged feral colonies.
Following the appearance of CCD in the fall of 2006, scientists began searching for its causes (see Rucker, Thurman, and Burgett, 2019). Bee scientists and regulators concluded early on that bees from CCD-afflicted colonies were infected with a broad range of known pathogens as well as with pathogens not reported before in the United States. Since these initial efforts, research has proceeded. Early speculation that cell phone signals may have been a cause of CCD were supplanted by alternative explanations with more longevity, including CCD being a new disease (possibly brought in by foreign bees), a response to malnutrition as a result of drought or habitat loss, resulting from exposure to stress (possibly induced by traveling long distances for pollination), or exposure to toxins and pesticides (in particular a class of insecticides called neonicotinoids that has seen increased use in recent years). A recent theme from the bee research community is that CCD is multifactorial and, as such, is not the result of a single causal agent.
In the past few years, some researchers have concluded that CCD is an overly broad label and have attributed higher winter mortality rates of the past decade to increasing resistance to treatments for Varroa and Nosema, to new strains of fungal parasites, and to the decreasing availability of forage for honey bees, in addition to the possible causal agents listed above.
Since the winter of 2006/2007, although the over-winter mortality rate of honey bees has increased substantially, U.S. colony numbers have grown. To make sense of this seeming incongruity, we briefly discuss aspects of honey bee biology, commercial beekeeping, and pollination markets. Our argument at the end of the discussion is that pollination markets function well and that commercial beekeepers have responded quickly and effectively to increased winter mortality rates. In the process, market responses have largely mitigated the impacts of CCD. Additional detail can be found in Rucker, Thurman, and Burgett (2012, 2019) and Burgett et al. (2010).
Honey bees collect nectar and pollen from flowering plants and, in the process of moving from flower to flower, enable plant reproduction. They are but one of thousands of animal species that pollinate about 90% of flowering plants, with the remaining 10% reproducing through pollination by wind and water. In the hive, bees transform the nectar into honey for later consumption (or extraction by beekeepers) and store the gathered pollen as a future protein source for the hive. Honey bees forage on almost anything that blooms, and this flexibility enhances their value to beekeepers.
A typical, full-strength colony of honey bees consists of a single queen and 25,000 to 40,000 worker bees. Queens usually live for about two years and, during that period, lay all the eggs in the hive. When a queen becomes less productive, the beekeeper replaces her with a newly fertilized queen, and the activities of the hive continue, largely uninterrupted. All the worker bees are sterile females with life spans of about six weeks in the summer. The colony also contains a small number of males, or drones, whose sole function is to mate with fledgling queens from other colonies.
Modern commercial beekeeping in the United States is highly migratory. Hives are moved by truck from crop to crop for pollination in the spring and, later in the year, to bee pasture for honey production. The U.S. crop that engages by far the most honey bees is almonds, all of which are grown in California. Rapidly expanding acreage in almond orchards, combined with rising almond pollination fees, have induced beekeepers from as far away as Florida and North Carolina to transport their colonies to California in the early spring, thereby further increasing the importance of migration. After early-season employment in almond orchards, bees are moved into fruit tree orchards and berry, melon, and vegetable fields to augment yield and fruit set.
Over the course of a season, individual worker bees die and the colony replaces them. At the super-organism level, some whole colonies die every year as well, with higher frequency in the winter. When a beekeeper walks into his bee yard and discovers a dead colony, he has several options for replacement. Surveys of Washington and Oregon beekeepers in the winters following the appearance of CCD suggest that the method used most frequently is “making increase” or “splitting” (see Burgett, Rucker, and Thurman, 2009; Caron et al., 2010; Caron and Sagili, 2011). The process requires the beekeeper to move approximately half of the brood and adult bees from a healthy hive to an empty hive. The new hives—known as nuclei colonies (or nucs, or splits)—require a fertilized queen. Newly mated queens are often purchased for this purpose from commercial queen breeders, who in aggregate produce and sell hundreds of thousands of queens per year. Following a successful split, the beekeeper will have two full-strength hives in about six weeks. The splitting process takes an experienced beekeeper about 20 minutes, and new queens bought in bulk by commercial beekeepers can be purchased for about $18. Beekeepers can split hives preemptively, in anticipation of future losses, or after colony loss early in the pollinating season.
Until the early twentieth century, beekeepers collected revenues primarily from honey sales. Pollination markets began to develop not long after the turn of the twentieth century. Few details are available regarding early pollination markets, but by the time Steven Cheung studied them in Washington state in the early 1970s, they were sufficiently well developed that pollination service providers could be located in the local yellow pages. (Yellow pages were listings of businesses found in large bound volumes known at the time as “telephone books.” See Rucker and Thurman (2010) for discussion and analysis of early U.S. pollination markets.) Cheung (1973) concludes that the 1970s pollination markets he investigated operated efficiently, and Rucker, Thurman, and Burgett (2012)–using more current and much more extensive data than Cheung–reinforce that conclusion.
Observed pollination fees result from the interactions among a variety of supply and demand factors (see Burgett, Rucker, and Thurman, 2010.) On the supply side, a primary determinant is the number of honey bee colonies managed by commercial beekeepers. The fact that U.S. colony numbers have fallen over time, from roughly 5.5 million in the mid-1960s to about 2.5 million in the early 2000s suggests that, ceteris paribus, the supply of pollination services has fallen (see Muth et al. 2003). However, the supply of pollination generated by a given number of colonies will also be affected by the price of honey; at the margin, beekeepers make trade-offs between honey production and providing pollination services. Other factors that have reduced the costs of commercial beekeeping include the development of the interstate highway system and flatbed trucks. Winter mortality rates also affect the supply of pollination services, although increased splitting of colonies in the late summer and fall can mitigate the reductions in colony numbers that accompany higher winter mortality rates.
Regarding the demand for pollination services, various crops employ bees, and they stock bees at different densities. Using data on Pacific Northwest beekeepers, Rucker, Thurman, and Burgett (2012) conclude that the average number of colonies used per acre ranges from less than 1.0 for cucumbers and squash and pumpkins to more than 2.0 for almonds and blueberries. The most important factor driving demand for bee pollination is almond production. Demand for pollination from almonds has increased dramatically over time as almond acreage has increased from 90,000 acres in 1950 to over 1,000,000 acres in 2018. Figure 3 displays the increase in almond pollination revenues between 1950 and 2016. The increase is due not only to the ten-fold increase in almond acres but also to increases in almond pollination fees, from roughly $1 per colony in 1950 to about $185 in 2016. As a result of this growth, in recent years, over 70% of U.S. colonies have been used to pollinate almonds in California in February. The tremendous importance of almonds in pollination markets is reflected in the fact that in 2016, 82% of all revenues from pollination services were from almonds (see Ferrier et al., 2018).
An important insight into the net effects of changing demand and supply conditions for pollination services over time can be seen by comparing past and present proportions of beekeeper incomes from honey and pollination services. In 1988, 11% of U.S. beekeeper revenue came from the provision of pollination services. By 2016, pollination revenues had increased to 41% of beekeeper revenues (Ferrier et al., 2018).
The preceding discussion suggests that (i) beekeepers respond to incentives transmitted through pollination markets and (ii) the relative and absolute magnitudes of the revenues from these markets have increased dramatically over time. Further, the increasingly migratory nature of commercial beekeeping is what allows markets to coordinate the narrow windows for providing pollination to crops that bloom only briefly, at different times each year, and in locations that span the United States.
Dramatically increased winter mortality rates over the last decade have been attributed to the appearance of Colony Collapse Disorder and in the past several years to a broader range of honey bee afflictions. Increased mortality has attracted unprecedented attention to the honey bee and the importance of the services it provides. Yet the number of honey bee colonies in the United States in recent years has risen, not fallen, even though colony numbers had fallen consistently for several decades prior to the appearance of CCD.
Why have colony numbers not fallen in concert with rising mortality rates? We conclude that the answer is found in an understanding of markets and the incentives they provide, combined with able deployment of beekeeping technology by beekeepers—factors overlooked by most researchers who investigate the seemingly precarious condition of honey bees. Beekeepers have always lost hives during the winter, and sustainable beekeeping requires them to replace dead and weak colonies using the splitting process and other methods. Since the onset of CCD, beekeepers have had to replace more hives to maintain colony numbers, and the time path of colony numbers shown in Figure 2 suggests they have succeeded in doing so.
The lesson we draw is that even in the face of a dramatic negative biological shock (i.e., the appearance of CCD and high mortality rates), well-functioning pollination markets combined with effective adaptation by commercial beekeepers have mitigated the impacts of the shock to such an extent that economic effects are hardly observable.
To conclude, consider the following questions that frame the public discussion and concern for bees, each followed by our response:
Question: Should we worry about the disappearance of managed honey bees?
Answer: No. Bees are managed livestock and beekeepers face appropriate incentives to manage honey bee disease problems.
Question: Should we worry that our food supply will be dramatically reduced?
Answer: No, and for similar reasons.
Question: Should we worry about the livelihoods of commercial beekeepers?
Answer: Costs of operating have increased as a result of CCD and other bee health problems. This causes beekeeper profits to fall. At roughly the same time, however, almond pollination fees have more than doubled, which makes beekeeping more profitable. Rucker, Thurman, and Burgett (2019) suggest that the latter effect outweighs the former, at least for commercial beekeepers whose bees pollinate almonds.
Question: Should we worry about the plight of nonmanaged pollinators?
Answer: From a biodiversity perspective, plausibly. Unlike honey bees, the services provided by unowned, nonmanaged pollinators are not transacted directly in markets and there are few market incentives promoting their conservation. It is useful to keep in mind here that data from managed honey bees have little direct relevance to the status or value of wild pollinators.
Rucker, R.R., W.N. Thurman, and M. Burgett. 2019. “Colony Collapse and the Consequences of Bee Disease: Market Adaptation to Environmental Change.” Journal of the Association of Environmental and Resource Economists. Available online: https://www.journals.uchicago.edu/doi/10.1086/704360
Bromenshenk, J.J., C.B. Henderson, C.H. Wick, M.F. Stanford, A.W. Zulich, R.E. Jabbour, S.V. Deshpande, P.E. McCubbin, R.A. Seccomb, P.M. Welch, T. Williams, D.R. Firth, E. Skowronski, M.M. Lehmann, S.L. Bilimoria, J. Gress, K.W. Wanner, and R.A. Cramer Jr. 2010. “Iridovirus and Microsporidian Linked to Honey Bee Colony Decline.” PloS ONE 5(10): e13181.
Burgett, M., S. Daberkow, R. Rucker, and W. Thurman. 2010. “U.S. Pollination Markets: Recent Changes and Historical Perspective.” American Bee Journal 150(1): 35–40
Burgett, M., R. Rucker, and W. Thurman. 2009. “Honey Bee Colony Mortality in the Pacific Northwest (USA).” American Bee Journal 149(6): 573–575.
Caron, D., M. Burgett, R. Rucker, and W. Thurman. 2010. “Honey Bee Colony Mortality in the Pacific Northwest: Winter 2008/2009.” American Bee Journal 150(3): 265–269.
Caron, D., and R. Sagili. 2011. “Honey Bee Colony Mortality in the Pacific Northwest: Winter 2009/2010.” American Bee Journal 151(1): 73–76.
Champetier, A., D.A. Sumner, and J.E. Wilen. 2015. “The Bioeconomics of Honey Bees and Pollination.” Environmental and Resource Economics 60(1): 143–164.
Cheung, S.N.S. “The Fable of the Bees: An Economic Investigation.” Journal of Law and Economics 16(1973): 11–33.
Cornman, R.S., D.R. Tarpy, Y. Chen, L. Jeffreys, D. Lopez, J.S. Pettis, D. vanEngelsdorp, and J.D. Evans. 2012. “Pathogen Webs in Collapsing Honey Bee Colonies.” PLoS ONE 7(8): e43562.
Ferrier, P.M., R.R. Rucker, W.N. Thurman, and M. Burgett. 2018. Economic Effects and Responses to Changes in Honey Bee Health. Washington, DC: U.S. Department of Agriculture, Economic Research Service, Economic Research Report ERR-246, March.
Mussen, E. 2007, October 17. “BSI: The Case of the Disappearing Bees.” Public Lecture, University of California-Davis. Summary article and video available online: http://169.237.77.3/news/dssericmussen.html [accessed January 27, 2015].
Muth, M.K., R.R. Rucker, W.N. Thurman, and C.-T. Chuang. 2003. “The Fable of the Bees Revisited: Causes and Consequences of the U.S. Honey Program.” Journal of Law and Economics 46: 479–516.
Muth, M.K. and W.N. Thurman. 1995. “Why Support the Price of Honey,” Choices 10(2): 19–23.
Nordhaus, H. 2011. The Beekeeper’s Lament: How One Man and Half A Billion Honey Bees Help Feed America. New York, NY: HarperCollins Publishers.
Oliver, R. 2019. “The Enigma of Nosema. Part 2.” American Bee Journal 159: 777–781.
Pernal, S.F. 2008. CAPA Statement on Honey Bee Losses in Canada (Spring 2008) – Final Revision. Canadian Association of Professional Apiculturists.
Pollan, Michael. 2007. “Our Decrepit Food Factories,” New York Times, December 16.
Robinson, W.S., R. Nowogrodzki, and R.A. Morse. 1989. “The Value of Honey Bees as Pollinators of U.S. Crops: Part I.” American Bee Journal 139(6): 411–423.
———. 1989. “The Value of Honey Bees as Pollinators of U.S. Crops: Part II.” American Bee Journal 139(7): 477–487.
Rucker, R.R. and W.N. Thurman. 2010. “Birds Do It, Bees Do It: Let’s Specialize and Exchange.” Presented paper at PERC workshop, “Ecosystem Services,” at the University of North Carolina-Chapel Hill.
Rucker, R.R., W.N. Thurman, and M. Burgett. 2012. “Honey Bee Pollination Markets and the Internalization of Reciprocal Benefits,” American Journal of Agricultural Economics 94(2012): 956–977.
Rucker, R.R., W.N. Thurman, and M. Burgett. 2019. “Colony Collapse and the Consequences of Bee Disease; Market Adaptation to Environmental Change. “ Journal of the Association of Environmental and Resource Economists 6(2019): 927–960.
Stipp, D. 2007, August 28. “As Bees Go Missing, a $9.3B Crisis Lurks.” Fortune. Available online: http://money.cnn.com/magazines/fortune/fortune_archive/2007/09/03/100202647/
Underwood, R.M., and D. vanEngelsdorp. 2007. “Colony Collapse Disorder: Have We Seen This Before?” Bee Culture 135(7): 12–15.
vanEngelsdorp D., J.D. Evans, C. Saegerman, C. Mullin, E. Haubruge, B.K. Nguyen, M. Frazier, J. Frazier, D. Cox-Foster, Y.Chen, R. Underwood, D. R. Tarpy, and J.S. Pettis. 2009. “Colony Collapse Disorder: A Descriptive Stud.” PLoS ONE 4(8): e6481.
vanEngelsdorp, D., R. Underwood, D. Caron, and J. Hayes. 2007. “An Estimate of Managed Colony Losses in the Winter of 2006-07: A Report Commissioned by the Apiary Inspectors of America.” American Bee Journal 147: 599–603.
White House Office of the Press Secretary. 2014, June 20. “Creating a Federal Strategy to Promote the Health of Honey Bees and Other Pollinators [Presidential Memorandum].” Available online: https://www.whitehouse.gov/the-press-office/2014/06/20/presidential-memorandum-creating-federal-strategy-promote-health-honey-b.

